
Хөдөө аж ахуйн их сургууль, “Монос- Фарм” ЭШТ, Монос дээд сургууль
Уламжлалт анагаах ухааны (УАУ) онол,хэрэглээний олон зуун жилийн туршлага дээр тулгуурлан ургамал амьтан, эрдсийн гаралтай нэгдлүүдээс дархлал сэргийлэх үйлдэлтэй шинэ эмийн бзлдмэл гарал үүслийн асуудал судлаачдын анхаарлыг ихээхэн (15) бөгөөд бид МУАУ д биеийн тамир тэнхээ сайжруулах зорилгоор хэрэглэгдэж ирсэн Шинэсэрхүү бударганы ургамалын дархлал сэргээх идэвхийг тогтоох .түүний идэвхи мембраны төлөвөөс хамаарах байдлыг нарийвчлан судлах зорилго тавьсан юм.
Судалгааны ажлын арга зүй:
Дархлалын урвалын эрчимд бэлдмэлийн үзүүлэх нөлөөг тодорхойлох арга зүй :
А.Бэлдмэлүүдийн шууд иммунзаслын идэвхийг 18-20 г жинтэй цагаан хулганад Т- хамааралт тест антиген болох хониныцусны улаан эсийн (ХЦуЭ) 10% -0.2 мл-р судсаар тарьж зэрэг үзүүлэлтээр харьцуулан тодорхойлов
Б. Дархлалын хурц дутагдалын үед дэлүүний эсийн үйл ажиллагаа өөрчлөгдөх баидал,түүнд бэлдмэлийн үзүүлэх нөлөөг тогтоохдоо цагаан хулганад СС1-4-ийг (40%-0.8 мл/кг 4 өдөр арьсан дор) тарьж “дархлал дутагдал”-ын загвар үүсгэн, 10% -0,2 мл ХЦУЭ судсаар тарьж, 5,7 дахь хоногуудад дэлүүний жин, дэлүүний индекс,дэлүүний эсийн тоо, гемагглютинины таньц зэрэг үзүүлэлтийг тодорхойлсон болно (А.И.Конопля1988, Л.Г.Прокопенко 1982).
В.“Дархлал дутагдал”-ын дараах янз бүрийн үе шатанд ӨХИП эрчимжих байдал, түүнд бэлдмэлийн хэрхэннөлөөлөх байдлыг дэлүүний эсийн мембранд агуулагдах малондиальдегид / МДА/, шиффийн суурийн /ШС/концентрацаар тодорхойлов (25).
-МДА-ыг тодорхойлохдоо 5-8%-ийн элэгний гомогенатад агуулагдах түүний концентрацийг тиобарбитурын хүчлийн тусламжтайгаар урвалд орж үүссэн триметины бүрдлийн концентрацаар Specol дээр 535 нм-т гэрлийн шингээлтийг авах замаар тодорхойлов (25).
-Шиффийн суурийн концентрацийг тодорхойлохдоо 5-8%-ийн элэгний гомогенатаас өөхний бодисыг “хлороформ + метанол”-аар хандлан ялгаж хлороформын давхаргад агуулагдах өөхний бодист ШС-ийн концентрацийг спектрофлоуриметр “Hitachi” дээр 350-360 нм-ийн өдөөлт, 430-440 нм-ийн гэрлийн шингээлтээр тодорхойлов (А.И.Стальная 1977).
Т. 3 лимфоцит эсийн тоо тодорхойлох арга зүй
Туршилт явуулсан төлгөн хонины цусан дахь В,Т лимфоцитын харьцангуй болон үнэлэхүй тоон үзүүлэлтийг А.М. Цымбал и др. (1983) нарын аргаар буюу Т-лимфоцитийг цусны улаан эстэй Е-РОК үүсгэх аргаар, В-лимфоцитийг хавсаргатай холбогдсон зимозантай наалдуулах урвалын аргаар тус тус тодорхойлов (19).
Фагоцито.зийн идэвх тодорхойлох арга:
Туршилтад авагдсан амьтдын (төлгөн хонины) макрофаг эсүүдийн фагоцитозын урвалын идэвхийг тэдний микроб залгих урвалын идэвхээр тодорхойлов (19).
Туршлагын амьдын эсийн мембранд альфа төлөбийн харьцангуй давамгаллыг өдөөхдөө фосфолипидийн 5мл/кг-аар тунгаар амаар уулгах аргыг хэрэглэн бетта төлөвийн давамгайллыг холестерин 250 мг /кг тунгаар уулгах аргаар тус тус сэдээж хавар намар зэрэг улирал хамааралтай бетта альфа төлөвийн давамгайлал бүхий улирлуудад ажиллатыг явуулав.
Шинэсэрхүү бударганы усан хандыг 0.15 гр/кг /хуурай бодис/ тунгаар уулгаж судалгааг гүйцэтгэлээ.
Судалгааны ажлын үр дүн:
А.Туршлагын амьтанд уусгэгдсэн дархлал дутагдлын эмгэг загварын уед Шинэсэрхуу бударганы бэлдмэлийн дархлал сэргээх идэвхийг судалсан дүн:
Бидний ажиглалтаар 18-20 г жинтэй цагаан хулганад GCL-4-ийн 40%-ийн тосон уусмалыг 0.8 мл/кг тунгаар 4 өдөр дараалан арьсан дор тарих үед эмчлээгүй хяиалт амьтдыг эрүүл амьтадтай харьцуулахад дэлүүний эсийн мембранд (ДЭМ) шиффийн суурийн хэмжээ 1.897 дахин нэмэгдсэн байгаа нь хяналт амьтдын дэлүүний эсийн мембранд CL*, СС1_3*-ийн прооксидант үйлдлийн нөлөөгөөр тосны ханаагүй хүчлүүд хэт исэлдэн задарч алдагдан, альфа шингэн төлөв эрс багасан, ханасан хүчил холестерин ихэсч, хатуу зуурамтгай бетта төлөв нэмэгдэх урьдчилсэн нөхцөл бий болсны илрэл юм (6,11,20). Өөхний хэт исэлдэлтийн процессын улмаас ДЭМ-д альфа:бетта төлөвийн хувийн жин ихэсч “бетта хамааралт” дархлал дутагдал явагдаж байгаагийн нотолгоо нь энэ үед хонины цусны улаан эсийн (ХЦУЭ) эсрэг явагдах дархлалын урвалын эрчим дэлүүний эсийн тоон үзүүлэлтээр 1.92 дахин буюу 47.9%-иар (Р<0.05), дэлүүний индексийн үзүүлэлтээр 1.36 дахин буюу 26,8%-иар (Р<0.05) буурч байгаа явдал юм. Харин Шинэсэрхүү бударганы усан хандны (ШБУХ, ХБОУХ) нөлөөгөөр мембрант CL*, ССL_3*-ийн оролцоотойгоор өдөөгдсөн өөхний хэт исэлдэлтийн процесс шиффийн суурийн тоон үзүүлэлтээр 1.83-2.0 дахин (Р<0.05) багасахын хамт дархлалын урвалын дээрх үзүүлэлтүүдийн хэмжээ 1.36-1.46 дахин ихэссэн нь түүний нөлөөгөөр мембранд ханаагүй фосфолипид, альфа шингэн төлөв алдагдаж, ханасан хүчил’ холестерин, бетта төлөв хэт давамгайлах үзэгдэл саатагдаж, бетта төлөв хамааралт дархлал дутагдлын процесс хориглогдох нөхцөл тавигдаж байгааг харуулж байна. Бидний энэ судалгаа нь гадаадын олон судлаачдын ажиглалттай нийцэж байгаа бөгөөд Ю.И.Губский (1989) нарын судалгаагаар ССL-ийг харханд 2 мл/кг тунгаар хэвлийд нэг удаа тарихад лимфоаит эсийн мембранд альфа төлөвийг болгодог ханаагүй хүчлийн хэмжээ 3 дахин багасч, үунтэй уялдан метоген бодисууд болох фитогемаглютний, конконавалины нөлөөгеер. Эсийн ДНХ-ийн бүтцэнд тимидин холбогдох хурд 2-8 дахин буурч ДНХ:РНХ-нйн 1.7 дахин багасч байсан бол иммуномодулятор үйлдэлтэй левамизолын нөлөөгөөр дархлалын эсийн мембранд агуулагдах ханаагүй хүчлийн хэмжээ нэмэгдэн митогены үйлдлийн дор тимидин холбогдох хурд ихэсч байгаа нь илэрчээ (6). Мөн Г.Б.Афонина (1991) нарын судалгаагаар дархлалын эсийн мембранд өөхний хэт исэлдэлтийн процесс эрчимжин сэдээгдэхийн хэрээр ханаагүй фосфолипид (фосфатмдилинозит, фосфатидилэтаноламин) улам багасч, ханасан фосфолипид (фосфатидилхолин, сфингомиелин) улам ихэсч Е- РОК, ЕАС-РОК, Fc-POK эсүүдийн тоо эрс цөөрөн лимфоцит эсийн ДНХ-д тимидин холбогдох хурд багасч, мембраны зуурамтгай чанар хэт нэмэгдэн мембрандхолбогдон байрласан рецептор, маркер, антигены хөдпөлгөөн 1.4 дахин суларч хемилюминесценцийн идэвх 1.3 дахин ихэсч байсан бол антиоксидант үйлдэлтэй аминдэм Е токоферолын нөлөөгөөр дархлалын эсийн мембранд хэт исэлдэлтийн процессын эрчим буурч, ханаагүй хүчил нэмэгдэн, мембраны хөдөлгөөн хурдасч, рецепторын экспресс сайжирч тимидины холбогдолт ихсэн “дархлал” сэргээх идэвхи ажиглагдсан байна.
Б, Шинэсэрхүү бударганы усан хандны иммунтроп үйлдлийн эрчим дархлалын эсийн мембраны төлөвийн өөрчлөлтөөс хамаарах байдал
Дархлалын урвалын эрчимжилт бууралт, антиген, митогены үйлдлийг хүлээн авах, хариу урвал үзүүлэх эрчим, иммуностимулятор, иммунотроп үйлдэлтэй эмт бодисын идэвх нь дархлалын эсийн мембраны төлөвөөс шууд хамаарахзүй тогтлын дагууд академич Р.В.Петров (1986) дархлалын урвалыг сэдээгч, өдөөгч үйлдэлтэй эмт бодис, вакцины химийн бүтцэнд дархлалын эсийн мембраны төлөвийг өөрчлөгч “модулятор” идэвхтэй химийн нэгдлийг заавал оруулж байх тухай шинэ санааг дэвшүүлсэн байдаг.Иймээр бид энэ удаагийн туршилтад мембраны бетта төлөвийн давамгайллыг сонгомлоор сэдээгддзг бодис болох холестершээр цагаан хулганы дархлалын эсийн мембрант хатуу зуурамтгай төлөвийг ихэсгэсэн болон мембраны альфа төлөвийн ихсэлтийг сонгомлоор сэдээгддэг бодис болох ханаагүй фосфолишдээр цагаан хулганы дархлалын эсийн мембранд шингэн урсамтгай төлөвийг зохиомлоор нэмэгдүүлсэн нөхцөлд иммуномостимулятор үйлдэлтэй Шинэсэрхүү бударгана (ШБ)-ы иммунотроп үйлдэл хэрхэн өөрчлөгддөгийг тогтоох зорилт тавьсан юм. Энд нэг зүйлийг зориуд анхааруулан хэлэхэд бид туршлагын цагаан хулганад дархлалын эсийн мембраны (ДЭМ) төлөвийн өөрчлөлтийг 3 үндсэн хувилбараар үүсгэв. Үүнд;
а. Дөрвөнхлорт нүүрстөрөгчийн оролцоотойгоор ДЭМ-д өөхний хэт исэлдэлтийн процессыг (ӨХИП) өдөөн альфа төлвийн хувийн жинг багасган, бетта төлөв хамааралт дархлал дутагдлыг үүсгэсэн буюу МБ-ийн дан бетта төлвийн давамгайлал бүхий /1983/ амьтад (мембраны дан бетта давамгайлал бүхий амьтад гэж цаашид нэрлэнэ).
б. Дөрвөнхлорт нүүрстөрегчийн оролцоотойгоор ДЭМ-д ӨХИП-ийг өдөөн бетта төлвийн давамгайлал сэдээхийн хамт холестерин уулгаж, МБ-ийн бетта төлвийн давамгайллыг давхар үүсгэсэн амьтад (мембраны бетта төлвийн давхар “бетта + бетта” давамгайлал бүхий амыад гэж цаашид нэрлэнэ).
в. Дөрвөнхлорт нүүрстөрөгчийн оролцоотойгоор ДЭМ-д ӨХИП-ийг өдөөн, мөн холестерин уулгаж бетта төлвийн давамгайлал сэдээхийн хамт улирлаас хамаарсан бетта төлөвийн давамгайллыг сэдээсэн мембраны бетта төлвийн гурвалсан давамгайлал бүхий “бетта + бетта + бетта” амьтад (цаашид мембран бетта төлвийн гурвалсан давамгайлал бүхий амьтад гэж нэрлэнэ).
Туршлагын цагаан хулганад CCL-4-ийг 40%-ийн 0.8 мл/кг тунгаар 4 өдөр дараалан арьсан дор тарьж, ДЭМ-д ӨХИП-г өдөөн, холестерин;фосфолипидын харьцаа буюу бетта төлвийг нэмэгдүүлсний дараа, холестерин 250 мл/ кг тунгаар амаар уулган,фосфолипид:холестерины харьцааг улам багасгаж мембраны бетта төлвийн давхар давамгайлая (СС14+холестерин) үүсгэсэн амьтдад (бетта + бетта) Шинэсэрхүү бударганыг хэрэглэх үед (СС1_4+холестерин + ШБ) хяналт амьтадтай (CCL4+ холестерин + ус) харьцуулахад дэлүүний эсийн тоо 1.5 дахин (Р<0.05), гемагглютинины таньц 1.9 дахин нэмэгдэж иммуномостимулятор үйлдэл илэрч байлаа.Нэг ижил туршилтын нөхцөлд зөвхөн мембраны төлвийг өөрчилсөн “СС14+ханаагүй фосфолипид” /бетта + альфа/ хувилбарын үед хяналт “СС1_4+ханаагүй фосфолипид + ус” амьтадтай / бетта + альфа + ус/ харьцуулахад туршилтын бүлгийн(CCL4+ ханаагүй фосфолипид + Шинэсэрхүү бударгана) амьтдад /бетта + альфа + ШБ/ дэлүүний эсийн тоо 1.62 дахин буюу 62.73%- иар гемагглютинины таньц 1.46 дахин буюу 46.75%- иар ихэсч Шинэсэрхүү бударганы усан хандны иммуномостимулятор үйлдэл илүү тод илэрч байгаа нь ажиглагдав. Ийнхүү Шинэсэрхүү бударганы усан ханд мембраны бетта төлөвийн давамгайллын үед ч(CCL4+.холестерин), альфа төлөвийг харьцангуй ихэсгэсэн нөхцөлд ч (CCL4 + ханаагүй фосфолипид) иммуномостимулятор үйлдэл үзүүлж байгаа боловч нарийн харьцуулалт хийх явцад мембраны альфа, бетта төлөвийн давамгайллаас шууд хамаарч Шинэсэрхүү бударганы (ШБ) усан хандны иммуномостимулятор үйлдлийн эрчимжилт харилцан адилгүй байгаа нь ажиглагдсан юм. Өөрөөр хэлбэл “CCL4+ холестерин + ШБ” буюу /бетта + бетта +ШБ/ хувилбарыг “CCL4+ ханаагүй фосфолипид +ШБ” хувилбартай харьцуулахад “CCL4 + ханаагүй фосфолипид +ШБМхувилбарын /бетта + альфа +ШБ/ үед дэлүүний эсийн тоо 1.68 дахин буюу 68.73%-иар (Р<0.05) нэмэгдсэн нь дэлүүнийэсийн мөмбраны альфа төлөвийн ихсэлтийн фон нь Шинэсэрхүү бударганы усан хандны иммуномостимулятор үйлдэл илрэх маш тохиромжтой урьдчилсан нөхцөл болохыг харуулж байна.Шинэсэрхүү бударганы усан хандны иммуномостимулятор үйлдлийн механизмыг дэлүүний эсийн мембраны түвшинд нарийвчлан тодруулах юм бол мембранд альфа төлөвийн давамгайлал нэмэгдэн холестерин.фосфолипидийн харьцаа багассан нөхцөлд (CCL4 + ханаагүй фосфолипид + ШБ) бэлдмэлийн антиоксидант үйлдэл хяналтын бүлгийн амьтадтай (CCL4+ ханаагүй фосфолипид + ус) харьцуулахад тод илэрч мембранд агуулагдах шиффийн суурийн хэмжээ 1.42 дахин буюу 29.8%-иар /хяналт амьтдад 1.6088Ю.16 ед, туршилтын амьтдад 1.1290±0.0522ед, Р<0.05/ багасч байсан бол мембраны бетта төлөвийн давхар давамгайлал сэдээгдсэн “CCL4+ холестерин + ШБГ туршилтын үед хяналтын амьтадтай (CCL4 +холестерин+ ус) харьцуулахад Шинэсэрхүү бударганы бэлдмэлийн антиоксидант идэвх харьцангуй суларч мембранд шиффийн суурийн хэмжээ дунд зэргийн эрчимтэйгээр багасан 11.4%иар буюу 1,12 дахин буурсан ерөнхий байдал илэрч байсан нь мембраны альфа төлөвийн давамгайллын үед Шинэсэрхүү бударганы усан хандны иммуномостимулятор үйлдэл хэт исэлдэлтийн процессын эрчим харыцангуй хадгалагдсан фонд дээр явагддагийг харуулж байна.
Хүснэгт 1. Шинэсэрхүү бударганы усан хандны иммуностимулятор үйлдэл улирал даган өөрчлөгдсөн мембраны- төлвөөс хамаарах нь
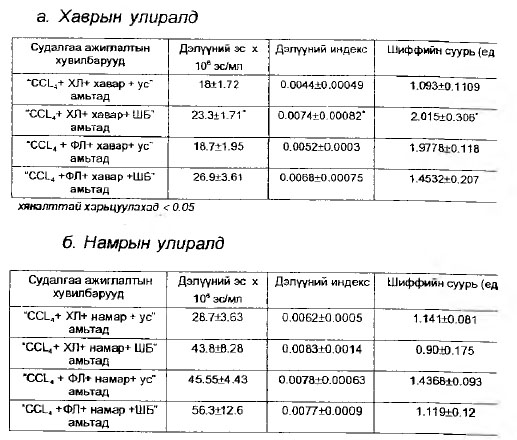
В.Шинэсэрхүү бударганы усан хандны иммуномостимулятор үйлдэл улирал дагаж өөрчлөгдсөн мембраны төлөвөөс хамаарах нь : Туршлагын цагаан хулганы дэлүүний эсийн мембранд хаврын улиралд мембраны бетта төлөвийн гурвал сан давамгайлал /бетта + бетта + бетта/ сэдээсэн үед “CCL4+ холестерин + хавар + ШБ” хувилбарыг хяналт амьтадтай (CCL4 + холестерин + хавар + ус) харьцуулахад дэлүүний эсийн тоо 1.3 дахин буюу 29.4%-иар /Р<0.05/, дэлүүний индекс 1.9 дахин /Р<0.02/, гемагглютинины таньц 1.88 дахин буюу 88.25%- иар /Р<0.05/ нэмэгдэж ШБ-ын усан хандны иммуномостимулятор идэвхи “бетта+бетта+бетта” нөхцөлд ч хадгалж байгаа боловч энэ хувилбарын үед уг бэлдмэлийн дархлал сэргээх идэвх бусад бүх хувилбарынхаас хамгийн сул түвшинд илэрч байгаа нь ажиглагдлаа. Хаврын улиралд туршлагын амьтдад ханаагүй тосны хүчил уулган, альфа төлөвийн хувийн жинг дээшлүүлсэн “CCL4+ хавар + ханаагүй тосны хүчил + ШБ” хувилбарын үед /бетта+бетта+альфа+ШБ/ хяналтын “CGL4 + хавар + ханаагүй фосфолипид + ус” амьтадтай I бетта+бетта-альфа+ус/ харьцуулахад дэлүүний эсийн тоо 1.43 дахин буюу 43.8%-иар (Р-<0.02)# дэлүүний индекс 1.3 дахин буюу ЗО./б^^-иар / ~ Р<0.05/, гемагшютинины таньц 163 дахин:^<0.02 нэмэгдэж иммуномостимулятор үйлдэл үзүүлэхийн xамт уг үйлдэл “CCL4 холестерин + хазар + ШБя-хувилбарын /бетта+бетта+бетта+ШБ/ үеийнхээс хүчтэй илэрч байгаа нь энэ үед Дэлүүний эсийн тоо 1,5 дахин (Р<0.05), гемагглютинины таньц 40%-иар буюу 1.4 дахин (Р<0.02) ихэссэнэзс харагдаж байна.
Намрын улиралд явуулсан “CCL4 + холестерин - + намар + ШБВ /бетта+бетта + альфа+ШБ/ туршилтын үед Шинэсэрхүү бударганы . иммуномостимулятор үйлдэл хяналтын “CCL4 + холестерин + намар + ус” амьтадтай / бетта+бетта+альфа+ус/ харьцуулахад дэлүүний эсийн тоо 1.52 дахин буюу 52.8%-иар Р<0.05, дэлүүний индекс 1.17 дахин буюу 17.7%-иар Р<0.01, гемагглютинины таньц 2.2 дахин /Р<0.05/,ихэссэн байдлаар илрэхийн хамт “CCL4 + холестерин + хавар + ШБ” хувилбартай / бетта+бетта+бетта+ШБ/ харьцуулахад дэлүүний эсийн тоо 1.88 дахин буюу 88.2%/ Р<0.05/ ихэсч альфа төлөвийн намрын улирал дагасан ихсэлт Шинэсэрхүү бударганы усан хандны иммуномостимулятор үйлдэлд мэдэгдэхүйц эерэг нөлөөлөл үзүүлж байлаа.
“Намрын улирал+ханаагүй тосны хүчил” гэсэн хувилбараар дархлалын эсийн мембранд альфа төлөв нэмэгдэх тутам Шинэсэрхүү бударганы иммуномостимулятор үйлдэл “антиоксидант” идэвхийн механизмаар илэрч байгаа нь “CCL4 + ханаагүй хүчил + намар + ШБ” буюу / бетта+альфа+альфа+ШБ/ хувилбарын үед хяналт амьтадтай харьцуулахад /CCL4 + ханаагүй фосфолипид + намар + ус/ дэлүүний эсийн мембранд шиффийн суурийн хэмжээ 1.28 дахин багасч байгаагаас харагдана(Хүснэгт № 1,2).Харин “CCL4 + холестерин + хавар + ШБ” хувилбарын /бетта+бетта+бетта+ШБ/ үед “CCL4 + холестерин + хавар + ус” хувилбартай харьцуулахад /бетта+бетта+бетта+ус/Шинэсэрхүү бударганы иммуностимулятор үйлдэл дэлүүний эсийн мембранд шиффийн суурийн хэмжээ харьцангуй ихэссэнтэй зэрэгцэн нөхцөлдөж явагдсан нь хатуу бетта төлөвийн хэт давамгайллын үед эмийн бодисын дархлал сэргээх үйлдэл сулавтар прооксидант үйлдлийн механизмаар буюу зуурамтгай хатуу төлөвийн “хүлээснээс” түр зуурын, зохицуулгатайгаар идэвхжсэн “хэт ислүүдийн” оролцоотойгоор чөлөөлөгдөн, салж идэвхижилтэнд орох хэлбэрээр явагддаг байж болохыг харуулж байна.
Г.Шинэсэрхүү бударганы усан хандны Т. В - лимфоцит эсти тоонд үзүүлэх нөлөө : Судалгаа.г эр хүйсний 19.8-22.7 кг жинтэй төлгөн дээр явуулсан бөгөөд Шинэсэрхүү бударгана /ШБ/ -ы усан хандыг 15%И3.5 г/кг тунгаар өдөрт 2 удаа , уулган эм уухаас өмнөх үеийг эм уусны дараах 3,4,14,21» 28 хоногуудтай харьцуулан Т-лимфоцит эсийн тоон тодорхойлолт явуулсан юм.Шинэсэрхүү бударганы усан хандны нөлөөгөөр Т-лимфоцит эсийн тоон үзүүлэлт уухын өмнөх үе буюу хяналтын үетэй харьцуулахад судалгааны 3 дахь хоногт 1.20 дахин буюу 20.52%-иар, судалгааний 5 дахь хоногт 1.31 дахин буюу 31.78%-иар, !Р<0.051 судалгааны 14 дэх хоногт 1.76 дахин буюу 76.15%-иар /Р<0.05/ , судалгааны 21 дэх хоногт 1,54 дахин буюу 54.96%-иар 14 /Р<0.05/, судалгаань! 28 дахь хоногт 1.98 дахин (Р0.01) ихсэн Т-лимфоцит эсийн тоог нэмэгдүүлж байлаа. Мөн В-лимфоцит эсийн тоо ч ийм хэлбэрээр нэмэгдэж байгаа нь ажиглагдсан юм.
Д. Шинэсэрхүү бударганы усан хандны макрофагийн эсийн залгих идэвхид үзүүлэх нөлөө :
Шинэсэрхүү бударганы усан хандны нөлөөгөөр макрофаг эсийн фагоцитозын идэвхи эм уухын өмнөх үе буюу хяналт үетэй харьцуулан тодорхойлоход судалгааны 5 дахь хоногт 1.23 дахин буюу 23.25%-иар /Р<0.05/,судалгааны 14 дэх хоногт 1.32 дахин буюу 32,09%-иар /Р<0.001/ , судалгааны 21 дэх хоногт 1.30 дахин буюу 30.93%-иар/ Р<0.05/, судалгааны 28 дахь хоногт 0. 37 дахин буюу 37.20%-иар ихсэн макрофаг эсийн фаГоцитозын идэвхи бэлдмэлийн нөлөөгөөр нэмэгдэж байлаа.
Эмнэл зүйн судалгааны зарим дүн :
Умайн хүзүүний хорт хавдар бүхий өвчтэй хүмүүс дээр явагдсан эмнэлзүйн судалгаанаас үзэхэд Шинэсэрхүү бударганаас гаргаж авсан “Салмон” эмээр иммуномониторинг явуулахад туяа эмчилгээний 30-35 грей, 70-75 грей авсан хяналтын эм хэрэглээгүй өвчтөнүүдтэй харьцуулахад Т супрессор эсийн тоо 1.2 багасч, үүнтэй зэрэгцэн нөхцөлдөж Тх/Тс-ийн харьцаа 1.21 нэмэгдэж, уг үйлдпийн идэвхи нь тималинаас 1.35 дахин өндөр байгаа нь тогтоогдлоо.Эцэст нь хэлэхэд бидний судалгаа нь МУАУ- нд янз бүрийн халдварт өвчинд болон биеийн тамир тэнхээ дээшлүүлэх зорилгоор хэрэглэдэг Шинэсэрхүү бударгана нь дархлал сэргээх идэвхи (ДСИ) үзүүлдэг, тэдгээрийн ДСИ нь мембраны альфа төлвийн давамгайллын үед эрчимжиж бетта төлвийн давамгайллын үед буурдгийг харуулахын хамт ямар ч эмийн бодисын тун хэмжээг зориуд нэмэгдүүлэлгүйгээр альфа төлвийн улирал хамааралт нэмэгдэлтийн биохэмт үзэгдэл хийгээд ханаагүй фосфолипид, липополисахарид, динитрофенол, Са+, К+, ионофор, деполяризатор зэрэг (Р.В.Петров, 1983) мембраномодулятор хүчин зүйлсийн оролцоотойгоор эзэн биеийн эффектор эсүүдийн мембраны төлвийг өөрчлөн, эмийн бодис, вакцин, эсрэгтөрөгч био бэлдмэлийн нөлөөллийг дээд зэргийн эрчимтэй хүлээн авах нөхцлийг бүрдүүлэн, үйлдлийг нэмэгдүүлэх (1,7,19,23) боломжтойг харуулж байгаа юм.
2. Афонина Г.Б., Бордонос В.Г., Роль свободнорадикального окисления мембранных липидов лимфоцмтов в развитии иммунологической недостаточности и её хоррекции токоферолом, “Иммунология”, 1990, №5, с. 33-36.
3. Богацкая Л.Н.3Кулъчицкий O.K., Потапенко Р.И. и др,, Липидный состав и свойства плазматических мембран при старении и некоторых видах эксприментальной патологии “Вестник АМН CCCF ”, 1990, №1, с.31-34.
4.Бородин Е.А.3 Побрецов Е.И., Карасевич Е.И. и др.3 Влияние выстраивания и выведения холестерина на вязкость липидного бислоя ш скорость окислительных процессов в мембранах микросом печени крыса, лБиохимияп,1981, №6, с. 1109-1119.
5.Бородин Е.А., Арчагов А.И., Теоретичеекое обсснование ислользования ненасыщенных липидов дпя восстановления структуры и функций гюврежденных биологических мембран, “Вестник АМН СССР”, 1985, №3, с. 84-90.
6. Пубский Ю.И., Афонина Г.В., Брюзгина Т.С, и др, Влияние левамизола на функциональную активность и жирнокмапотный состав лимфоцитов при ловреждении клеточных мембран тетрахлорметаном, “Вопр. Мед. химии”, 1989, М®2, с.64-68,
7. Крепс Е.М.,Липиды клеточных мембран, Л.Наука, 1981, с.340
8.Кузьмина И.Л., Стойда Л.В., Влияние холестерина на кооперативность Са-АТФ-азы саркоплазматического ретикулума скелетных мыщц кролика, “Блю.эсклер. биол. и мед.”, 1984, №12, с. 671-673.
9. Кузнецов В.А., Мелконян С.Г., Лисовская М.Л. и др. Изменение микровязкости плазматической мембраны и чувствмтельности к ПГЕ, тромбоцитов, 63 уйёбпу хбе 22 ё 40° С. “Волр. Мед. химии”, 1987, №4, с. 28-30.
10. Лопухин Ю.А., Азизова О.А., Маркин С.С. и др. Вторичный атеросклеротический иммунодефицит, “Вестик АМН СС-СР”, 1983, №33 с. 3-10.
11. Лопухин Ю.М., Владимиров Ю.А., Арчашв А.И., Холестериноз, М. Мед, 1983, с.352
12. Ляшенко В.А., Механизмы активациит уноле петентных клетск,у Медицина, 1988,
13. Панасенко О.М., Вольнова Т.В., АзизоваО.А. и -др, ПОЛ фактор, способствующий накоплению холестерина в -клетках при атерогенезе, “Бюл. экспер. биол. и мед.\" , 1988, №9 с. 277-280.
14. Панасенко О.М., Ли ХО Ик, Азизова О.А., Влияние холестерина на скорость восстановления спинных зондов в липосомах ионами, “Биол. мембраны”, 1988, №8, с. 872-878.
15. Хайдав Ц., Алтанцэцэг Б., Варламова Т.С., Лекарственные растения В Монгольской медицине, УБ, 1985,с. 390
16. Халилов Э.М., Ли B.C., Азизова О.А., Структурно-функциональный анализ мембран эритроцитов с различным содержанием холестерина. “Бюл.эскпер. биол. и мед.”, 1982, №1, с. 81-86
17. Хохлов А.П., Ярыгин К.Н.3 Бурлакова Е.Б., Синтетические фенольные антиоксиданты - полуфункциональные модуляторы биологичесКих мембран, “Биофизика”, 1989, №1, с. 133-143.
18. Цветких В.Е., Крылов., Лернер Г.Я. и др, Роль структурно- функциональных изменений клеточных мембран в формировании иммунного ответа у больных хроническим пиелонефритом, Тех. архив”, 1991, №2, с. 105-108.
19. Шарманов А.Т.,Влияние полиненасыщенных жирных кислот на иммунный ответ и окислительный метаболизм макрофагов, “Журнал микробиологии и эпидимилогической мммунологии”, 1985, №6, с. 110-111.
20. Binkova В. Lipid pereoxidation induced changes in physical properties of annular lipids in rat brain syn¬aptosomal membranes, Gen. physiol. Biophysic, 1990, Jun, 9(3), pp.311-318.
21.Brasitus T.A.,Davidson N.O.,Schachter D. Variations in Dietary triacylglycerol saturation alter the lipid composition and fluidity of rat intestinal plasma membranes, Biochimica et Biophysica Acta, 1985, vol.812, pp.460-472
22. Huber L.A.,Xu Q.B.,Jurgens G. Correlation of lymphocyte Lipid composition membrane microviscosity and mitogen response in the age. Eup. J. immunol, 1991,v.21,n 11, pp.2761-2765
23. Richard D.,Berlin and Judith P.Fera Changes membrane microviscosity associated with phagocy¬tosis effects of colchicine. Proc. Nath. Acad, Sci. USA, 1977,N 3, vol. 74.
24.Shinitzky M.3 Inbar M. Difference in Microviscosity induced by Different Cholesterdl Lev¬els in the Surface Membrane Lipid Layer of Normal Lymphocytes and Malignant Lymphoma Cells, J.Mol, Biol., 1974, v.85, pp. 603-615.
25.Tappel A.L. In vitro Lipid pereoxydation, Free radicals in Biology, 1979, pp. 3-15


