
АУИС-ийн Хүн Судлалын Үндэсний Төв, Япон Улсын Улаан Загалмайн Нийгэмлэгийн Цус Судлалын Төв
201 Khalkha-Mongolians living in central part of Mongolia were typed for DRB1 locus using PCR-MPH with further confirmation by SSCP, and PCR-RFLP method . Gene frequencies were calculated by maximum-likelihood method using the program developed for the Xlth International Histocompatibility Workshop. A total of 35 different DRB! alleles were identified in this group of samples. The most frequent DRB1 allele was DRB1*0301(11.1%), followed by DRB1*0701(9.7%) and DRB1*1101(8.5%), while the most common phenotypes were
DRB1*1501DRB1*1101(4.61%), DRB1*0301DRB1*0401(4.6 1 %), DRB1*0301DRB1*1101 (4.61%) and DRB1*0301DRB1*0701 (4.61%). One novel DRB1 allele and two rare types, DRB1*1111 and DRB1*1504, recently described in Jews and the Dai minority of China, respectively, were identified in Mongolians. The sequence of the novel allele, found in Mongolians was identical with that of DRB1*1403 except for the fact that it had codon for phenylalanine instead of the codon for leucine at aminoacid position 67. The name DRB 1* 1427 has been officially assigned to the novel allele by the WHO Nomenclature Committee for Factors of the HLA System. Data on the nucleotide sequence have been reported to GenBank and may be obtained under the accession number D86504.
Pp. 6-10, Tables 3, Picture 1, References 13
Энэхүү судалгаандаа бид Монголын хүн амын доторхи HLA-ситемийн II ангийн генүүдийн тархалтын зүй тогтлыг судлах зорилго тавьсан юм.
Судалгааны материал, арга зүй:
1. Судалгааны материал. Судалгаанд уугуул монголчуудыг Монгол Улсын хүн амын үлэмж олонхи болох халх ястнаар төлөөлүүлсэн бөгөөд Өвөрхангай,Архангай, Төв, Булган, Говь-Алтай, Баянхонгор, Завхан аймгуудын хүн амаас цуглуулсан нийт 201 сорьцод HLA-DRB1 локусын генүүдийг молекул аргаар тогтоосон болно.
2. ДНХ ялгах арга, Геномын ДНХ-г хөлдөөж хадгалсан 1 мл цуснаас Герман Улсад үйлдвэрлэсэн ДНХ ялгах цомгийг зааврын дагуу ашиглан ялгасан болно (QIAmp, QIAGEN, Germany).
3. ПГУ-МПЭ (Полимеразын Гинжин Урвал -Ми кропланшет и йи Эрлийзжүүлэлт)
ПГУ-МПЭ урвалыг Kawai нарын аргаар явуулсан ба (Kawai et a!., 1994 a, 1994b). урвалд ашигласан праймеруудыг 5-р хүснэгтэнд толилуулав. ПГУ-г35 циклтэй явуулсан бөгөөд цикл тус бүр 94°С-т 30 секунд (денатурацийн шат) 55°С-т 30 секунд (праймер холбогдох шат) 72°С-т 1 минут (ДНХ олшрох шат) шатуудаас бүрдэж байв.
Биотиноор тэмдэглэгдсэн ПГУ-ын бүтээгдэхүүнийг микропланшетийн ёроолд бэхжүүлсэн зондтой 58°С-т эрлийзжүүлэлт явуулсан ба үлдэгдэл ПГУ-ын бүтээгдэхүүнийг ТМАС (тетраметкпаммониум хлорид) уусмалаар автомат угаагчаар 3 дахин угаасан болно. Зондтой холбогдсон ПГУ-ын бүтээгдэхүүн авидинтэй холбогдсон пероксидазтай өнгийн урвалд орох бөгөөдтүүнийг 415 nrn гэрлийн долгионд хэмжсэн болно (зураг 1).

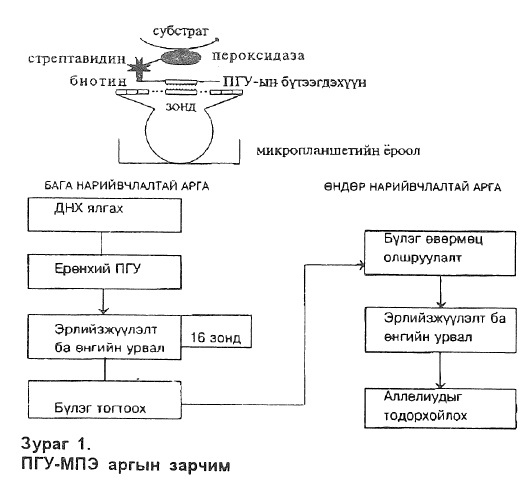
DRBl-инн еронхнй тодорхойлолтонд 16 зонд ашигласан бөгөөд өндөр нарийвчлалтай DRl-DRBl-нйн тодорхойлолтонд 5 зонд DR2-DRBl-ийн тодорхойлолтонд 9 зонд DR4-DRB 1-ийн тодорхорхойлолтонд 12 зонд, DR12/8-DRB1-ийн тодорхойлолтонд 10, DR3/11/6-DRB1-iihh бүлгийн тодорхойлолтонд 12 зонд тус тус ашигласан болно. (зураг 2).
4. ПГУ-РФУП (Полимеразын Гинжин Урвал-Рестрикцийн Фрагментийн Уртын Политорфизм)
DRB142 дэд бүлгийн аллелиудыг тодорхойлоход П ГУ-РФУП аргыг ашигласан болно. 20тл ПГУ-ыы уусмалнь300 ng ДНХ, 1.5 мМ MgCL2, 50mMTris-HCL (рН 8,3), 10mМ аммонийн сульфат, деоксирибонуклеознд трифосфат тус бүрээс 12.5 тМ, ираймер тус бүрээс lOpmol болон 0.5 U Taq полимераза фермент агуулж байв. Урьдчилсан денатурацийг 94°С-т 5 мшгут явуулсны дараа ПГУ нь 94°С-т 1 минут, 64°С-т 1 мннут бүхий 35 циклэос бүрдэж байв. Эцсийн инкубацийг 5 минутын хугацаатай72 °С –т явуулсан болно. DR1*1201 болон DRBP1202 аллелиудыг салгаж тодорхойлохын тулд Fok 1 рестрикцийн ферментийг ашигласан болно.
5. ПГУ-ДХБП (Полимеразын Гинжин Урвал-Дан Хэлхээний Байгууламжийн Полиморфизм)
DRB1 аллелиудыг эцсийн байдлаар баталгаажуулахын тулд ПГУ-МПЭ-ийн өндөр нарийвчлалтай урвалын дараа ПГУ-ДХБП аргыг Bannai нарын хувилбараар ашигласан болно (Bannai etal., 1994). 10мл ПГУ-ын уусмал нь 30 ng ДНХ, lmM MgCl2, 50тМ Tris-HCl (рН 8,8), 10мл аммонийн сульфат ба деоксирибонуклеознд трнфосфат тус бүрээс 20 мл, праймер тус бүрээс 0.5 mM-ийг болон 0.2U Taq полимераза фермент агуулж байв. ПГУ-ыг 95°С-т 5 минут байлгасны дараа цикл тус бүр 95°С-т 1 минут денатурацийн шат, 53°С-62°С-т 1 минут праймер холбох болон ДНХ олшруулах шатуудаас бүрдэл болсон нийт 30 цикл явуулсан ба эцсийн инкубацийг 72°С-т 5 минут байлгав.
Олшруулсан ДНХ-г 95°С-т денатураци явуулах уусмалд (95% формалимнд, 20mM EDTA, 0~05% бромфенол хөх, 0.05% ксилен цианол FF) 5 минут байлгаад мөсөн дээр гэнэт хөргөөх замаар дан хэлхээ үүсгэж электрофорезнйг 12.5%-ын полиакриламидын гельд 4°С-ын тогтмол температурт явуулав.
- Нуклеотидуудын дараалал тодорхойлох Харьцуулалтыы стандартДНХ байгаагүй гурван тохиолдолд тухайн аллелийг нуклеотидуудын дарааллыг тогтоох замаар тодорхойлсон болно. Нуклеотндуудын дарааллыг дидеокснор хэлхээг битүүлэх аргаар (Sanger et al, 1977) автомат ДНХ секвенаторын тусламжтай (373А, Applied Biosystems, USA) шууд секвенс хийж тодорхойлов.
- Тооцооны аргууд
Генийн давтамжуудыг хамгийн үнэнд дөхүүлсэн (maximum likelihood) аргаар HLA- судлалын XI Ажлын Зөвлөгөөнөөр Т. Imanishi-ийн зохиосон тооцоолон бодох программыг ашиглан тогтоосон ба энэхүү арга нь нэгдүгээрт судалгаанд хамрагдсан бүх локусууд кодоминант хэлбэрээр удамшдаг, хоёрдугаарт локус бүрт зовхон blank аллелиуд рецессив хэлбэрээр удамшдаг болох, гуравдугаарт нэгж популяциуд Харди-Вайнбергийн тэнвэрт байдалд байгаа гзсэн гурван болзолд тулгуурладаг юм (Imanishi etal., 1992b).
Судалгааны үр дүн:
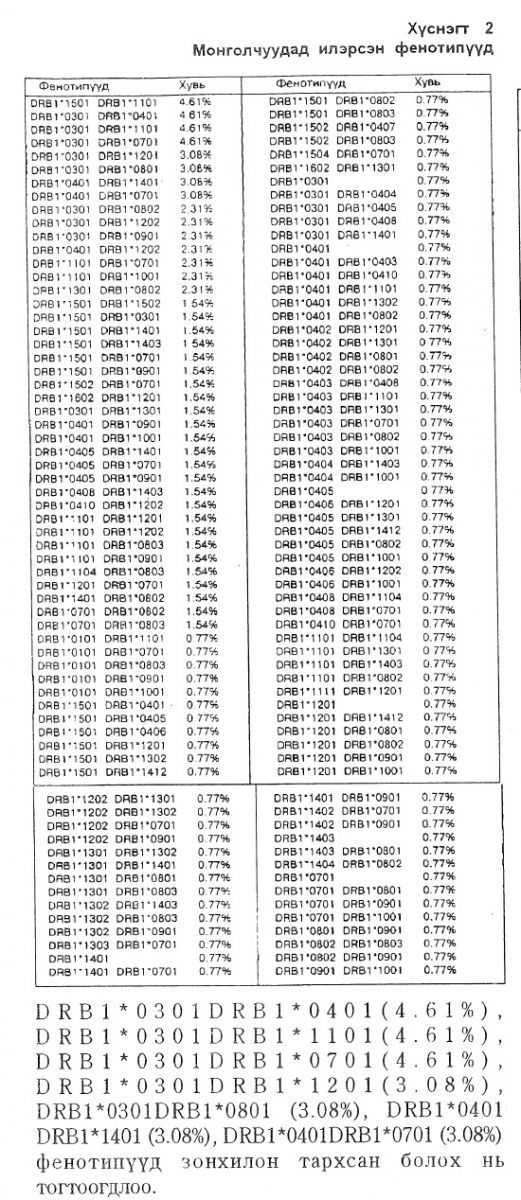
Фенотигүүдийн тархалт. HLA-DRB1 локусып хүрээнд монголчуудад нийт 130 фенотип илэрснийг хамгнйн өндөр хувьтайгаас нь дарааллуулан 2-р хүснэгтэнд үзүүлэв. Бидний судалгаагаар монголчуудад DRB1*1501DRB1*1101 (4.61%.), фенотипууд зонхилон тархсан болох нь тогтоогдлоо.
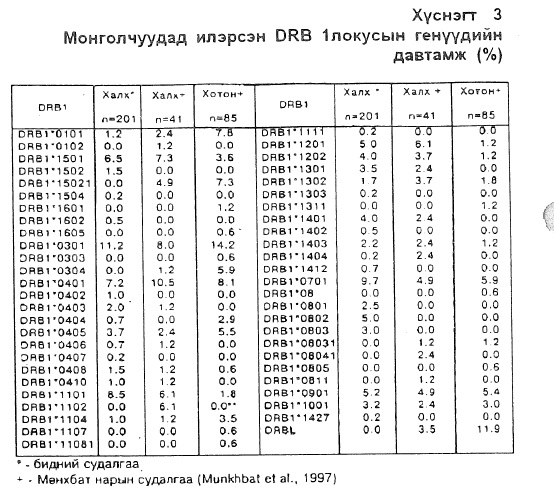
Генүүдийн тархалт . DRB1 локусын хүрээнд халх монголчуудад бидний судалгаагаар нийт 35 аллель ген илэрсэн бөгөөд тодорхой серологийн өвөрмөц чанарт бататгах аллелиудыг нэгэн бүлэгт хамруулж бүлэг тус бүрийн хүрээнд илэрсэн генүүдийг доорхи байдлаар тодорхойлов (хүснэгт 3). Үүнд: DR1 бүлэгт зөвхөн DRB 1*0101 (1,2%) аллель ген илэрсэн байхад DR2 бүлэгт DRB14501, DRB14502, DRB14602 болон DRB1M504 аллелнуд иlэрснээс DRB14501 (6,5%) аллель хамгийн өндөр давтамжтай байна. DR3 бүлэгт ганц DRB 1 *0303 (11,2%) аллель ген илэрсэн ба тэрбээр DRB1 локусын хүрээнд монголчуудын дунд хамгийн өндөр давтамжтай илэрсэн аллель ген юм. DR4 бүлэгт 9 аллель илэрсэнээс DRB 1*0401 (7,2%), DRB1*0405 (3,7%), DRB1*Q403 (2,0%), DRB1*0408(1,5%), DRB1*0402 (1,0%), DRB1*0410(1,0%), DRB1*04Q4 (0,7%), DRBl*040fi (0,7%), DRB 1*0407(0,2%) аллель генүүд багтаж байна. DR5 бүлэгт 5 аллель илэрсэн ба DRB1*1101 (8,5%) аллель ген хамгийн өндөр давтамжтай байгаа ба DRB1*1201 (5,0%) ген нь тухайн бүлэг дотроо удаах түгээмэл давтамж бүхий ген болох нь тогтоогдсон байна. DR6 бүлэгт хамгийн өндөр давтамжтайгаар DRB 1*1401 аллель ген илэрсэн бөгөөд DRB1*1301(3,5%), DRB1*1401 (2,2%), DRB1*1302 (1,7%) аллелиудмөнөндөр давтамжгай илэрсэн байхад DRB1*1303 (0,2%), DRB1*1404 (0,2%), DRB1*1402 (0,5%), DRB1*1412 (0,7%) аллель генүүд давтамж багатай илэрсэн байна. DR14 дэд бүлэгт монголчуудын дунд нэгэн шинэ аллель ген илэрсэн явдал бидний судалгааны нэн сонирхолтой үр дүн болсон билээ. DR7 бүлгийн DRB1*0701 (9,7%) аллель нь DRB1 локусын хүрээнд монголчуудьш дунд DRB 1 *0301-ийн дараахь өндер давтамжтай ген болох нь тогтоогдлоо. DR3 бүлэгт 3 янзын аллель ген илэрсэн байна: DRB 1*0802 (5,0%), DRB1*0803 (3,0%), DRB1*0801 (2,5%) зэрэг генүүд болно. DR9 бүлгийн DRB1*0901 аллель нь 21 (5,2%) тохиолдолдВЕЮ бүлгийн DRB1*1001 аллель нь 13 (3,2%) тохиолдолд тус тус илэрсэн болно (Chimge etal, 1997).
Бид өөрсдийн судалгааныхаа дүнг хотончууд болон халхуудын дунд HLA-системийн II ангийн генүүдийн молекул төвшинд хийгдсэн Мөнхбат нарын судалгааны дүнтэй харьцуулж үзэхэд (MunkhbatetaL, 1997) HLA-DRB1 локусын хүрээнд DRB1*1111, DRB1*1303, DRB1*1504, DRB1*1602, DRB1*1412, DRB1*0402, DRB1*0404, DRB1*0407, DRB1*0801, DRB1*0802, DRB1*0803, DRB1*1402, DRB 11427 аллелиуд буюу нийт 13 аллель ген бидний судалгаагаар халхуудад илэрсэн хэрнээ Мөнхбат нарын судалгаагаар илрээгүй, нөгөө талаас DRB1*0102, DRB1*15021, DRB1*0304, DRB1*1102, DRB1*08031, DRB1*08041, DRB1*0811 зэрэг нийт 7 аллель ген бидний судалгаагаар илэрсэн ба дээрхи судалгаагаар халхуудад илрээгүй зүй тогтол ажиглагдаж байна. Харин DRB1*1601, DRB1*1605, DRB1*0303, DRB1*1107, DRB1*11081, DRB1*1311, DRB1*0805 зэрэг нийт 8 аллель ген хотончуудад илэрсэн байхад халх ястанд огт илрээгүй онцлог ажиглагдлаа.
HLA-DRB1 аллелиудын молекул түвшинд хийгдсэн бидний судалгааны явцад гурван тохиолдолд ямар аллель ген болох нь тодорхой бус байдал илэрсэн ба эдгээр аллелиудын нуклеотидуудын дарааллыг шууд секвенсийн аргаар тогтооход анхеврейчүүдэд тодорхойлогдож HLA-системийн нэршилд шинээр бүртгэгдсэн DR5 бүлгийн DRB1M111 (Smith AG et al., 1994), мөн түүнчлэн Хятадын Дан ястнаас анх шинээр илрүүлсэи DR2 бүлгийн DRB1*1504 аллель (Fan et al., 1994) зэрэг сүүлийн үед нээгдсэн нэн ховор генүүд болох нь илэрсэн ба харин гуравдахь тохиолдол нь урьд өмнө бүртгэгдэж байгаагүй нэгэн цоо шинэ аллель ген болох нь тогтоогдсон болно.
Шинэ DRB1 1427 аллель генийн судалгаа. Бидний судалгаагаар илэрсэн шинэ аллель ген нь DR6 бүлгийн DR14 дэд бүлэгт хамаарагдах бөгөөд энэхүү шинэ аллель генийн' –(DRB1 * 1427) шууд секвенсийн аргаар тогтоосон нуклеотидуудын дарааллыг DRB1 локусыы бусад аллелиудын дараалалтай харьцуулж үзэхэд DRB 1*1427 нь DRB 1*1403 аллелиас 67-р байрлал дахь амин хүчлийн кодоны эхний нуклеотид болох цитозин тимин болж өөрчлөгдснөөр ялгарч байв (зураг 2). Ийнхүү ганц нуклеотидьн өөрчлөлтөөс улбаалан DRB 1*1403 аллелийн 67-р байрлал дахь лейцин (СТС) ф(Ш1шаланин (ТТС) болж хувирсан байна. Иймд DRB 1*1427 хэмээх энэхүү шинэ аллель нь DRB1*1403 аллель генээс цэгийн мутацийн үр дүнд үүссэн байж болзошгүй боловч нөгөө талаас HLA-системийн асар өндөр полиморфизм нь п) л төлев тогтворжуулах шалгарал, генийн конверси болон генийн дрейф зэрэг хүчин зүйлсийнхар^шцан үйлччэлийн үр дүнд оршин тогтдог гэж үздэг (Ohta, 1992; Nei and Hughes, 1992; Parham, 1992) учир шинэ аллель үүсэхэд 67- дахь байрлалдТТС кодон агуулсан аль нэг аллель болон DRB1* 1403 аллелиудын хоороыд энэхүү хэсэгт генийн конверси явагдсан нь илүү магадлалтай болов уу (Kashiwaseetal., 1997).
Шинэ аллелийг ДЭМБ-ын Нэршлийн комиссоос 1996 оны 6р сард албан ёсоор хүлээн зевшөерч HLA-системийн нэрш1шд HLA-DRB1* 1427 нэрээр бү])тгэсэн бөгөед тухайн генийн 2-р экзоны бүтэн секвенс нь D86504 дугаараар GenBank-ийн GENOMENET пуклеотидуудын мэдээлчийн санд хадга.лагдаж байгаа болно.

Дүгнэлт
Монгол Улсын хүн амын үндсэн төлөөлөгч болох халх ястны хүн амд НLA системийн II ангийн DRB1 локусын полиморфизмийн судалгаанаас доорх дүгнэлтүүд гарч байна. Үүнд:
1. Монголчуүдып дунд тус локусын хүрээнд нийт 130 фенотип илэрснээс DRB1*1501DRB1*1101 (4.61%) >
DRB1*0301DRB1*0401 (4Л>1%) > DRB1*0301DRB1*1101 (4.61%) > DRB 1*0301 DRB 1*0701 (4.61%) DRB1*0301DRB1*1201 (3.08%) > D R В 1 * 0 3 0 1 D R В 1 * 0 8 0 1 ( 3 . 0 8 % ) > D R В 1 * 0 4 0 1 D R В 1 * 1 4 0 1 ( 3 . 0 8 %) > DRB1*0401DRB1*0701 (3.08%) фенотипүүд зонхилон тархсан байна.
2.Уг локусын хүрээнд нийт 35 DRB 1 аллель ген илэрсэн бөгөөд DRB1*0301(11.1%), DRB1*0701(9.7%), DRB1*1101(8.4%) генүүд харьцангуй өндөр давтамжтай тохиолдлоо.
3. Монголчуудын дунд бидний судалгаагаар DR5 бүлэгт анх еврейчүүдэд илэрсэн DRB1*1111; DR2 бүлэгт Хятадын Даи үндэстэнд анх тодорхойлогдсон DRB1*1504; DR14 бүлэгт япончуудад анхлан тогтоогдсон DRB 1*1412 зэрэг сүүлийн үед нэршилд шинээр бүртгэгдсэн нэн ховор аллелнуд илэрсэн явдал монголчуудын генийн сан нэн өргөн дотоод генетик хувьсамжтай болохыг гэрчилж байна.
4. HLA-системийн DRB1 локусын DR14 дэд бүлэгт монголчуудын дунд нэгэн шинэ аллель ген илэрсэн бөгөөд ДЭМБ-ын дэргэдэх HLA-системийн Нэршлийн Комиссоос тэрбээр HLA-системийн албан ёсны нэршилд DRB 1*1427 гэсэн нэрээр бүртгэгдсэн явдал бидний судалгааны нэн сонирхолтой үр дүн боллоо.
Discrimination of human HLA-DRB1 alleles by PCR-SSCP (single strand conformation polymorphism) method. Eur JImmunogenet 1994:21:1-9.
2.Chimge N, Tanaka H, Kashiwase K, et al. The HLA-System in the Mongolian population. Tissue An¬tigens, 1997:49:5:477-483.
3.Fan LA, Smith AG, Chandanayingyong D and Hansen JA. DRB1*1504 (DR2Dai): Anew DR2 allele identified in the Dai minority population of Southeast China. Tissue Antigens 1994:44:326-328.
4.ImanishiT, AkazaT, Kimura A, Tokunaga K and Gojobori T. Estimation of allele and haplotype frequencies for HLA and complement loci. In: Tsuji K et al. eds. H1A 1991, Vol.l, Oxford University Press, 1992:76-79.
5.Kashiwase K, Chimge N, Kawaga Y, et al. Anew HLA-DR14 allele, DRB1*1427, identified in the Mon¬golian population. Tissue Antigens, 1997:50:682-684.
6.Kawai S, Maekawajiri S, Tokunaga K, Juji T and Yamane A. A simple method of HLA-DRB typing using enzymatically amplified DNA and immobilized probes on microtiter plate. Hum /жтшЫ.1994а:41:121-126.
7.Kawai S, Maekawajiri S, Tokunaga K, et al. HLA-DRB typing using PCR-MPH (microtiter plate-hybridization). MHCIRS 1 (Suppl.):1994:121-125.
8.Munkhbat B, Sato T, Hagihara M, et al. Molecular analysis of HLA polymorphisms in Ююton-Mongolians. Tissue Antigens, 1997:50:124-134.
9.Nei M and Hughes AL Balanced polymorphism and evolution by the birth-and-death process in the MHC loci. In: Tsuji K et al. eds. HLA 1991, Vol.l, Oxford University Press, 1992:27-38.
10.OhtaT. Diversifying selection, gene conversion,and random drift interactive effects on polymorphism at MHC loci. In: Tsuji K et al. eds. HLA 1991, Vol.1,
Oxford University Press, 1992:20-27.
11. Parham P. Evolution of class I HLA polymorphism: selection and drift. In:TsujiKet al. eds. HLA 1991, Vol.l, Oxford University Press, 1992:72¬82.
12.Sanger F, Nicklen S and Coulson AR. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 1977:74:5436-5467.
13.Smith AG, Safirman C, Kelso C, Amar A, HansenJAand Brautbar C. Two new DR52 associated DRB1 alleles, DRB1*1111 and*1312, identified by PSR/SSOP and confirmed by DNA sequence. Tissue Antigens 1994:44:52-56.


